Aspect psychologique de nos horloges biologiques
La chronobiologie
Le champ de recherche que l’on nomme aujourd’hui la chronobiologie s’intéresse aux rythmes biologiques c’est-à-dire à la manière dont notre organisme génère ses oscillations physiologiques et garde ses divers systèmes synchronisés. À partir des premières constatations de l’existence de cycles plus ou moins longs dans le corps humain, nos connaissances sur la chrono-biologie se sont rapidement complexifiées.
On s’est ainsi rendu compte que l’alternance du jour et de la nuit n’est pas la source de nos rythmes circadiens, mais qu’elle ne fait que synchroniser une véritable horloge biologique autonome. Cette horloge centrale qui reçoit la synchronisation de la lumière du jour est située dans le noyau suprachiasmatique.
On s’est aussi rendu compte de l’importance du phénomène à travers la plupart des grands systèmes du corps. Peu importe la variable physiologique que l’on mesurait, métabolisme cellulaire, tempé-rature corporelle ou sécrétion de nos différentes hormones, tout obéissait à des cycles avec des maximums et des minimums qui leur sont propres.
D’autres paramètres physiologiques connaissent des fluctuations endogènes importantes au cours de la journée. C’est le cas de la sécrétion de plusieurs hormones. La mélatonine, fabriquée dans la glande pinéale, est presque indécelable dans le sang pendant la journée. Elle commence à être sécrétée en milieu de soirée, à mesure que la lumière diminue, et atteint son pic de sécrétion entre 2 heures et 4 heures du matin.
Parmi toutes les études qui nous ont permis d’appréhender les articulations subtiles entre ces différents rythmes, l’apport des expériences en isolement temporel est des plus considé-rables. Plusieurs chercheurs se sont en effet demandé ce qu’il adviendrait si on isolait complètement une personne des indices habituels de l’alternance des jours et des nuits (lumière, bruits, etc.) ? Allait-elle continuer à s’endormir à son heure habituelle ou bien observerait-on un avancement ou un recul de son cycle d’activité ?
Ces expériences en isolement temporel ont été effectuées à plusieurs reprises, les premières fois dans des grottes, endroits à température constante naturellement isolés de tout. La première grande confirmation a été que les rythmes circadiens persistaient malgré l’isolement, ce qui prouvait que nous avons tous dans le cerveau une « horloge endogène ».
Sauf que ces expériences montraient aussi que cette horloge n’était pas parfaite et retardait de quelques minutes à chaque jour. En d’autres termes, notre rythme circadien naturel était un peu plus long que 24 h, soit de 24,2 à 25,5 heures, selon les études. Ça peut sembler petit, mais cela signifie qu’un cycle de 24,5 h transfor-merait en seulement 3 semaines l’activité diurne d’un individu en activité nocturne !
Ces expériences d’isolement temporel, ne datent pas d’hier. Déjà en 1938, Nathaniel Kleitman et son collègue Bruce Richardson avaient passé 32 jours sans indices temporels dans une grotte du Kentucky, aux États-Unis. En 1962, le français Michel Siffre passe quant à lui 2 mois dans un glacier souterrain des Alpes-Maritime en France. Siffre, qui avait 23 ans lors de cette expé-rience, allait par la suite retour-ner deux autres fois sous terre pour mesurer l’influence de l’absence de repères temporels sur ses rythmes biologiques à différents âges. La troisième fois, en 2000, il était âgé de 61 ans et est resté 73 jours en l’absence de tout repère temporel.
Un des phénomènes les plus spectaculaires observé durant ces expériences
d’isolement temporel est le décalage qui s’opère par rapport au cycle
jour/nuit à l’extérieur du laboratoire ou de la grotte. Mais aussitôt
l’expérience terminée, la personne resynchronise en quelques jours son cycle
d’éveil et de sommeil sur celui du jour et de la nuit, tel qu’illustré sur le
diagramme ci-dessous. Celui-ci montre un enregistrement quotidien du
cycle veille-sommeil d’une personne pendant un mois et demi. Chaque ligne
horizontale est un jour; les traits continus marquent les phases de sommeil, et
les lignes pointillées, les phases d’éveil. Le triangle représenté dans chaque
cas marque le point de la journée où la température corporelle était la plus
basse.
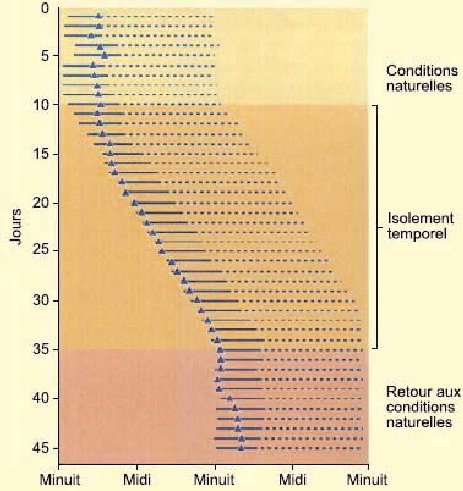
Lors de cette expérience, des enregistrements contrôles ont d’abord été fait pendant 9 jours dans des conditions naturelles de variation d’éclairage et de bruits qui accompagnent la journée. Puis, pendant 25 jours, tous ces repères ont été coupés pour laisser la personne évoluer selon son rythme endogène. Dans ces conditions, le cycle veille-sommeil se maintient mais a tendance à s’allonger à environ 25 heures. Après plusieurs semaines d’isolement, ces cycles peuvent s’allonger pour atteindre 30 à 36 heures. Le sujet peut par exemple rester éveillé durant 20 heures, puis dormir ensuite pendant 12 heures et se sentir tout à fait bien ainsi. (Source : d’après Dement, 1976)
Autre phénomène intéressant durant ces expériences : le point minima de température corporelle se déplace de la période de fin de sommeil, vers le début du sommeil. On assiste donc à une désynchronisation des cycles comportementaux (veille-sommeil) et des cycles physiologiques comme la température du corps. C’est cette désynchronisation qui serait à l’origine des problèmes causés par le décalage horaire. Enfin, durant les onze derniers jours de l’expérience illustrée sur le diagramme, les conditions initiales ont été réintroduites : la longueur du cycle se normalise alors avec les conditions d’éclairement jour-nuit, et la température corporelle se retrouve à son minimum vers la fin des périodes de sommeil.
Aspect psychologique de nos horloges biologiques
Les structures cérébrales qui nous réveillent et nous endorment
L’activation corticale nécessaire à l’éveil est rendue possible par un « réseau exécutif de l’éveil » comprenant l’hypothalamus postérieur, le thalamus intra-laminaires et le télencéphale basal. Mais ces structures sont elles-mêmes sous l’influence d’un réseau assez complexe d’une dizaine de structures situées en cascade du tronc cérébral inférieur au télencéphale basal et qui prennent en charge l’éveil.
Très schématiquement, les composantes de ces systèmes modulateurs ascendants peuvent être groupées en deux grandes voies qui ont toutes deux leur origine dans une partie du noyau réticulé du bulbe rachidien.
- La voie ventrale se projette vers l’hypothalamus postérieur et le noyau de Meynert du télencéphale basal (neurones à acétylcholine) : c’est la voie réticulo-hypothalamo-corticale.
- La voie dorsale active les noyaux mésopontins cholinergiques, la formation réticulée mésencéphalique (neurones à asparte/glutamate) et le thalamus : c’est la voie réticulo-thalamo-corticale.
En plus d’être à l’origine de l’éveil, plusieurs des noyaux de ces deux voies utilisant l’acétylcholine et le glutamate comme neurotransmetteur sont en partie responsables de l’activation corticale du sommeil paradoxal.
Parmi les noyaux les plus importants du tronc cérébral impliqués dans le système d’éveil, on retrouve donc :
- La formation réticulée mésencéphalique, qui se projette massivement sur les noyaux thalamiques, qui vont ensuite influencer tout le cortex. Son rôle en est un de désynchronisateur du cortex au sens large, favorisant l’éveil mais aussi le sommeil paradoxal. Il s’agit de l’ancien « système réticulé activateur ascendant » qui est maintenant considérée comme une simple partie du réseau de l’éveil.
- Les noyaux mésopontins cholinergiques, qui se projettent également sur le thalamus. L’acétylcholine produite par ces noyaux exerce une double action : elle diminue l’activité du noyau réticulaire thalamique appartenant au système du sommeil ; et elle active les neurones thalamocorticaux impliqués dans l’éveil.
- Les noyaux réticulés bulbaires magnocellulaires, dont les neurones sont cholinergiques ou à asparte/glutamate, est à l’origine à la fois de la voie réticulo-thalamo-corticale et de la voie réticulo-hypothalamo-corticale. Ses projections vont donc sur la formation réticulée mésencéphalique et les noyaux mésopontins cholinergiques ainsi que sur le télécenphale basal et l’hypothalamus postérieur.
- Les noyaux du locus coeruleus, situés dans la partie dorsale du pont, et dont les projections noradrénergiques influencent des structures cérébrales comme le thalamus, l’hippocampe et le cortex. L’activité du locus coeruleus est maximale chez le sujet éveillé et actif, réduite durant un éveil calme, encore plus réduite en sommeil lent, et complètement abolie en sommeil paradoxal.
- Les noyaux sérotoninergiques du raphé antérieur (ou supérieur), qui projettent de la sérotonine vers l’hypothalamus et le cortex. Actifs pendant l’éveil, l’effet global des noyaux du raphé antérieur est éveillant et, contrairement aux autres groupes aminergiques, leur lésion non seulement n’entraîne pas de somnolence, même passagère, mais provoque une insomnie prolongée de plusieurs jours. Cette apparente contradiction viendrait du fait que ce système, qui innerve l’hypothalamus antérieur au niveau de l’aire préoptique et l’horloge circadienne située dans le noyau suprachiasmatique, semble mesurer la durée et l’intensité de l’éveil. L’éveil pourrait ainsi provoquer éventuellement sa propre inhibition par rétroaction négative. En d’autres termes, le fait d’être éveillé durant un certain temps conduit au sommeil.
L’ensemble de ces structures du tronc cérébral reçoit des collatérales des afférences sensorielles et végétatives qui participent ainsi au maintien de leur activité (voir encadré). Il s’agit d’un réseau complexe dont l’excitation pharmacologique d’un élément amène l’activation de tous les autres. Cette organisation redondante explique aussi pourquoi l’inactivation d’un seul système est suivie, après quelques jours, d’une récupération complète de l’éveil. Aucune des structures décrites, prise isolément, n’est donc indispensable à l’activation corticale.
Le cerveau durant le sommeil paradoxal
Le sommeil paradoxal, avec son EEG semblable à celui de l’éveil, est le produit de l’interaction complexe entre des noyaux du tronc cérébral, des structures du système limbique et des aires corticales. D’autres de ses caractéristiques étonnantes ont aussi des mécanismes qui ont pu être localisés. C’est le cas des mouvements oculaires rapides qui surviennent durant le sommeil paradoxal et dont on ignore la fonction. Ils sont produits par la formation réticulaire pontique et transmis aux couches motrices des colliculi supérieurs. Les neurones colliculaires projettent à leur tour sur la formation réticulaire pontique paramédiane (FRPP) qui coordonne la durée et la direction des mouvements oculaires. Autre caractéristique singulière du sommeil paradoxal dont on a localisé la source : la paralysie quasi-totale du corps qui l’accompagne.
L’activité nerveuse intense enregistrée durant le sommeil paradoxal excite en effet la grande majorité des neurones corticaux, y compris ceux du cortex moteur primaire. Ces neurones moteurs génèrent ainsi des séquences d’activité organisées commandant des mouvements du corps mais seul les muscles respiratoires, oculaires, ceux de l’oreille interne pourront actualiser la commande motrice. Celle-ci ne parviendra jamais jusqu’aux motoneurones des membres.
Durant le sommeil paradoxal, l’augmentation d’activité de neurones cholinergiques de la protubérance va exciter d’autres neurones de la formation réticulée pontique qui utilisent le glutamate comme neurotransmetteur. Ceux-ci vont à leur tour envoyer des projections vers le bulbe rachidien où ils vont activer des interneurones qui relâchent de la glycine. Ce sont ces neurones bulbaires du noyau magnocellulaire, dont les axones descendent dans la moelle épinière, qui vont inhiber fortement les motoneurones en les hyperpolarisant.
Quant aux pointes ponto-géniculo-occipitales (PGO) survenant de façon intermittente durant le sommeil paradoxal, elles seraient elles aussi issues de la formation réticulée pontique. Elles se propageraient ensuite jusqu’au cortex occipital par l’entremise du thalamus bien qu’elles soient, comme leur nom l’indique, plus aisément détectables dans les relais du système visuel que sont les corps genouillés latéraux.
Les ondes PGO font partie des différents événements phasiques du sommeil paradoxal qui accompagnent les mouvements oculaires rapides, les changements dans le rythme respiratoire et cardiaque. Elles peuvent être générées en l’absence de sommeil paradoxal en stimulant la protubérance avec de l’acétylcholine, en particulier dans la région péribrachiale. C’est dans cette région réticulaire, autour du pédoncule cérébelleux supérieur et sous le locus coeruleus, que sont générées les ondes PGO.
Plusieurs de ces neurones qui se projettent sur le thalamus sont cholinergiques. Ils font une brève décharge de potentiels d’action juste avant chaque onde PGO du côté ipsilatéral.
La sérotonine du système du raphé inhibe les ondes PGO en hyperpolarisant ses cellules génératrices. On comprend alors pourquoi l’arrêt de l’activité des cellules sérotoninergiques lors du passage du sommeil lent au sommeil paradoxal déclenche les ondes PGO.